酵母单杂、酵母双杂、酵母三杂,仅仅一个字的区别,你对它们了解吗?这些经常用到的实验,它们的原理你确定都搞清楚了吗?如果没有,那么今天你就来对地方了,因为伯远生物(https://plant.biorun.com)下面打算非常详细的给你讲一遍它们的原理,让你以后不再为不懂原理而烦恼!
在了解酵母杂交实验之前我们先来了解一个转录因子——Gal4,了解它可以帮助大家更好的理解酵母杂交实验的原理!
Gal4转录因子
真核基因转录受大量调控因子调节。进化保守的Gal4是真核细胞普遍存在的转录激活因子。在半乳糖的诱导下,酵母Gal4结合在半乳糖代谢酶系GAL基因启动子的上游激活序列(Upstream activating sequence,UAS)上,调控Galactokinase(Gal 1),Galactose permease(Gal2),Galactose 1-phosphate uridyltransferase(Gal7),Galactose mutarotase and UDP-galactose 4-epimerase(Gal10)等基因的转录。
酵母单杂交与酵母双杂交
酵母单双杂实验的意义
生物体内各种生物分子之间的复杂调控网络构成了生命过程的基础。这些网络控制着胚胎如何发育成多细胞生物、该生物如何维持稳态以及如何对病原体或疾病作出反应等。参与这些网络的生物分子包括DNA、RNA、蛋白质和代谢产物等小分子。这些生物分子之间一般会存在相互作用共同调控复杂的生命过程,为了解析这些相互作用,酵母单杂、酵母双杂就成为了必不可少的研究手段!
酵母单杂原理
酵母单杂(Y1H)是在酵母双杂的基础上发展而来的,按理说应该先介绍酵母双杂的原理,但是考虑到酵母双杂还存在核体系与膜体系的区别,这里就先介绍酵母单杂吧!
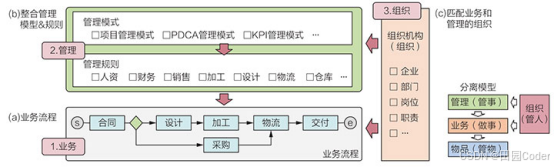
基本的Y1H分析包括两个载体(图1):(1)包含感兴趣的DNA(通常被称为诱饵(Bait))和报告基因融合的载体,该报告基因编码的蛋白易于检测;(2)包含感兴趣的转录因子(TF,通常被称为猎物(Prey))和酵母转录激活域(AD)融合的载体。这两种载体都被转入到合适的酵母菌株中,如果TF在酵母核的环境中与DNA结合,那么AD就会诱导报告蛋白的表达。重要的是,无论TF是激活因子还是抑制因子,酵母AD都会激活报告因子,因此,Y1H分析的是蛋白质和DNA之间的相互作用。
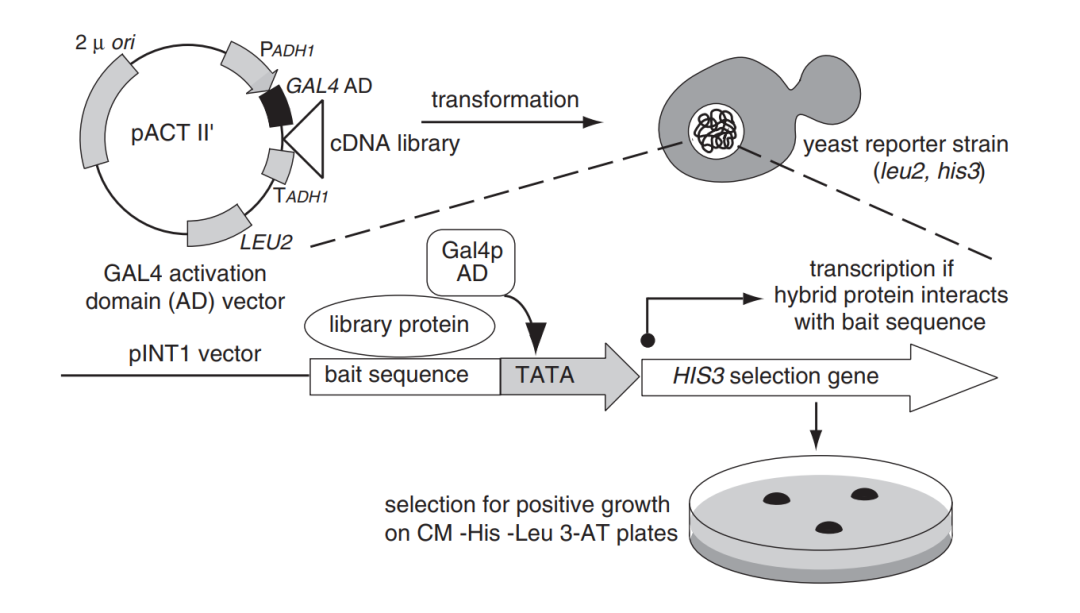
图1 酵母单杂实验原理(Ouwerkerk, Pieter BF.and Annemarie H., 2001)(不同的实验体系可能会存在一些差异,但原理基本都差不多,这里主要帮助大家理解原理)。
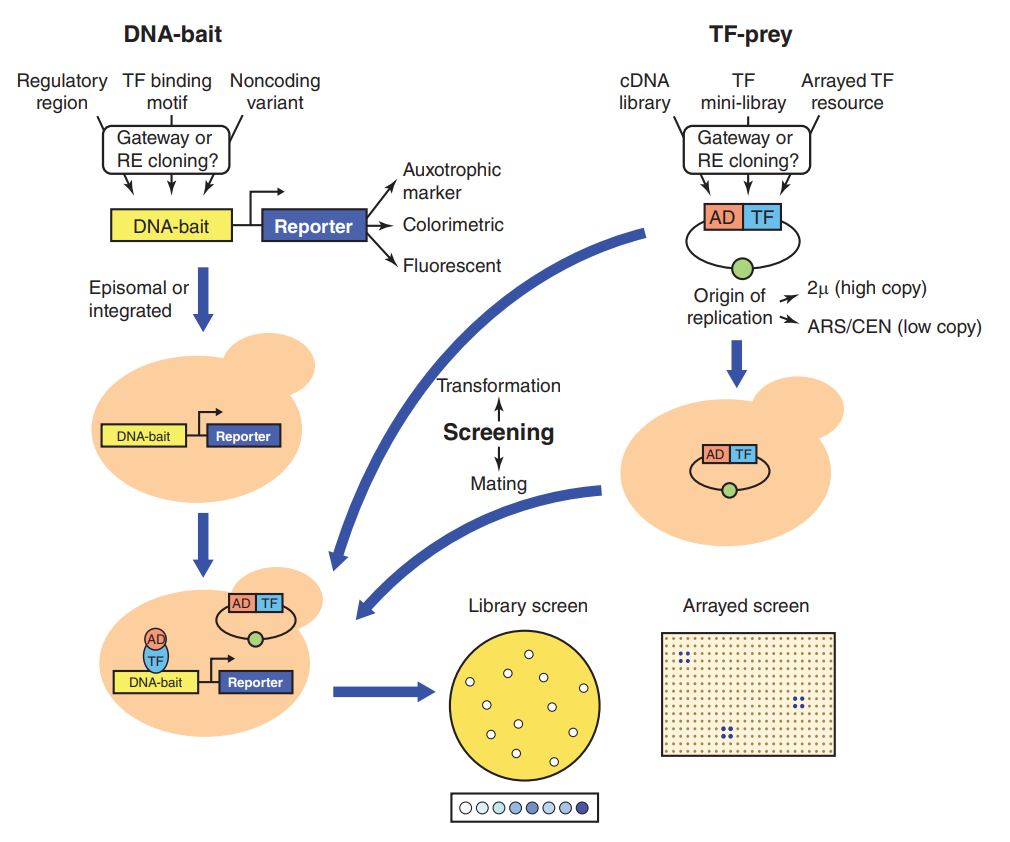
图2 Y1H分析的变化和注意事项概述(Sewell Jared A.and Juan I. Fuxman Bass, 2018)。利用限制性内切酶(RE)或Gateway技术将复杂调控区、转录因子结合基序或非编码变异的DNA序列克隆至报告基因上游。这些结构最常被整合至酵母基因组中以产生DNA诱饵菌株。然后将DNA诱饵菌株转化或与一组猎物克隆(ORF融合到Gal4的激活结构域AD上)交配。编码猎物的载体可以是低拷贝(ARS/CEN)或高拷贝(2μ)的。筛选可以以文库进行筛选,其中阳性菌落必须进行测序和重新测试,同时,也可以以阵列的形式进行筛选,阳性菌落的位置表明相互作用的转录因子(TF)的身份。
说明:酵母单杂筛库是以DNA为诱饵筛选与其互作的转录因子,如果你想要通过转录因子筛选与其互作的DNA,用到的方法应该是ChIP-seq。这个大家一定要区分清楚哦!
酵母双杂原理
(1)核体系酵母双杂
1989年,Fields和Song通过描述一个遗传系统来检测酵母细胞(Fields Stanley and Ok-kyu Song,1989)中蛋白质-蛋白质的直接相互作用,从而彻底改变了蛋白质相互作用分析的研究方法。在此之前,两种蛋白质之间的相互作用主要是利用生化技术来进行研究的。这一全新的分析工具的发现是由真核转录因子的分子分析引发的。在该项发现被公布的前几年,Ptashne实验室发现了酵母中转录激活因子Gal4的模块化结构。他们表明,Gal4结合一个特定的DNA序列(上游激活序列,UAS),从而在半乳糖存在的情况下激活转录。如果分离成两个片段,N端片段仍然可以与DNA结合,但在半乳糖存在时不激活转录,而激活的功能是由C端片段(Keegan et al., 1986)介导的。然而,这两个片段可以相互作用和非共价重建成一个有功能的Gal4。因此,研究者确定了Gal4的两个不同功能域:N端DNA结合域(DBD)和C端DNA激活域(AD),这两个独立的功能域独立于另一个的存在而保持各自的功能。
受这些发现的启发,Fields和Song利用转录因子Gal4的模块化特性来监测蛋白质之间的相互作用。其基本思想是将感兴趣的两个蛋白X和Y分别融合到Gal4的DBD和AD上,这样X和Y之间的相互作用可以重建Gal4转录因子的功能,然后驱动报告基因的表达(图1)。在第一个诱饵结构中,蛋白质X(如葡萄糖传感器SNF1)被融合到含有DBD的GAL4的N端部分(GAL4DBD)。在第二个猎物结构中,猎物蛋白Y(如调控蛋白SNF4)被融合到含有AD的Gal4的C端部分(GAL4AD)。融合蛋白在酵母中的表达以及诱饵和猎物之间的相互作用让Gal4两个分开的功能域重组成一个功能性的Gal4转录因子。然后Gal4招募RNA聚合酶II,引发GAL1-lacZ融合基因的转录和表达。lacZ报告基因编码半乳糖苷酶,可使用比色底物标记酵母细胞进行检测(Fields Stanley and Ok-kyu Song,1989)。
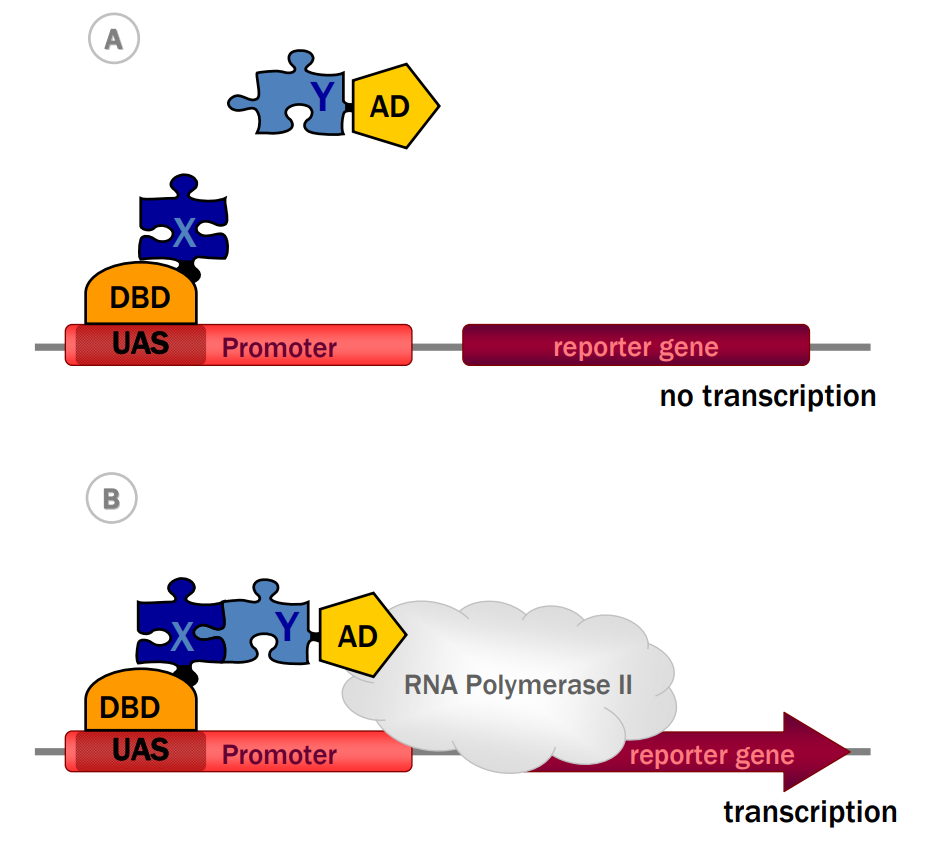
图3 酵母双杂实验原理(Brückner et al., 2009)。(A)感兴趣的蛋白质X,被融合到DNA结合域(DBD),该结构被称为诱饵。潜在的相互作用蛋白Y被融合到激活域(AD)上,该结构被称为猎物。(B)诱饵,即DBD-X融合蛋白,结合启动子的上游激活子序列(UAS)。诱饵与猎物(即AD-Y融合蛋白)的相互作用,招募AD,从而重建一个功能性转录因子,导致RNA聚合酶II的进一步招募和报告基因的后续转录。
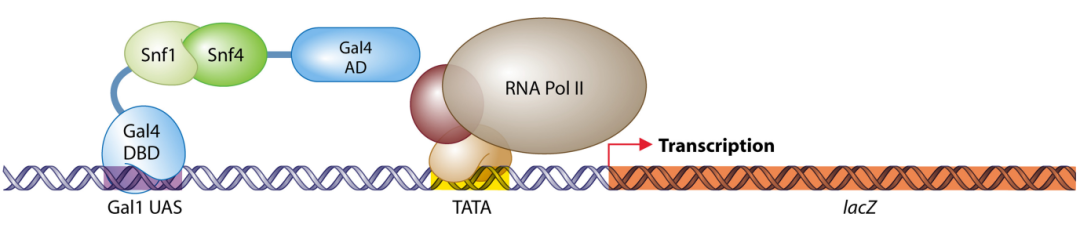
图4 蛋白SNF1与SNF4互作示意图(Stynen et al., 2012) 。
核体系酵母双杂的衍生应用
应用一
自从酵母双杂交技术开发以来,就被证明是检测蛋白质相互作用的有力工具。在另一方面,有机配体的二聚体可以导致与受体融合的两个独立蛋白质的二聚或寡聚。为了使酵母双杂交系统能够用于研究小分子配体与受体的相互作用,研究者设想使用两个不同的小有机配体合成异源二聚体作为第三个杂化分子,使DNA结合结构域与其中一个配体的受体融合,激活结构域与第二个配体的受体融合,从而激活报告基因。该系统被称之为酵母三杂交系统,可以用于直接识别编码感兴趣的配体的受体cDNA,或筛选与特定受体结合的新配体。
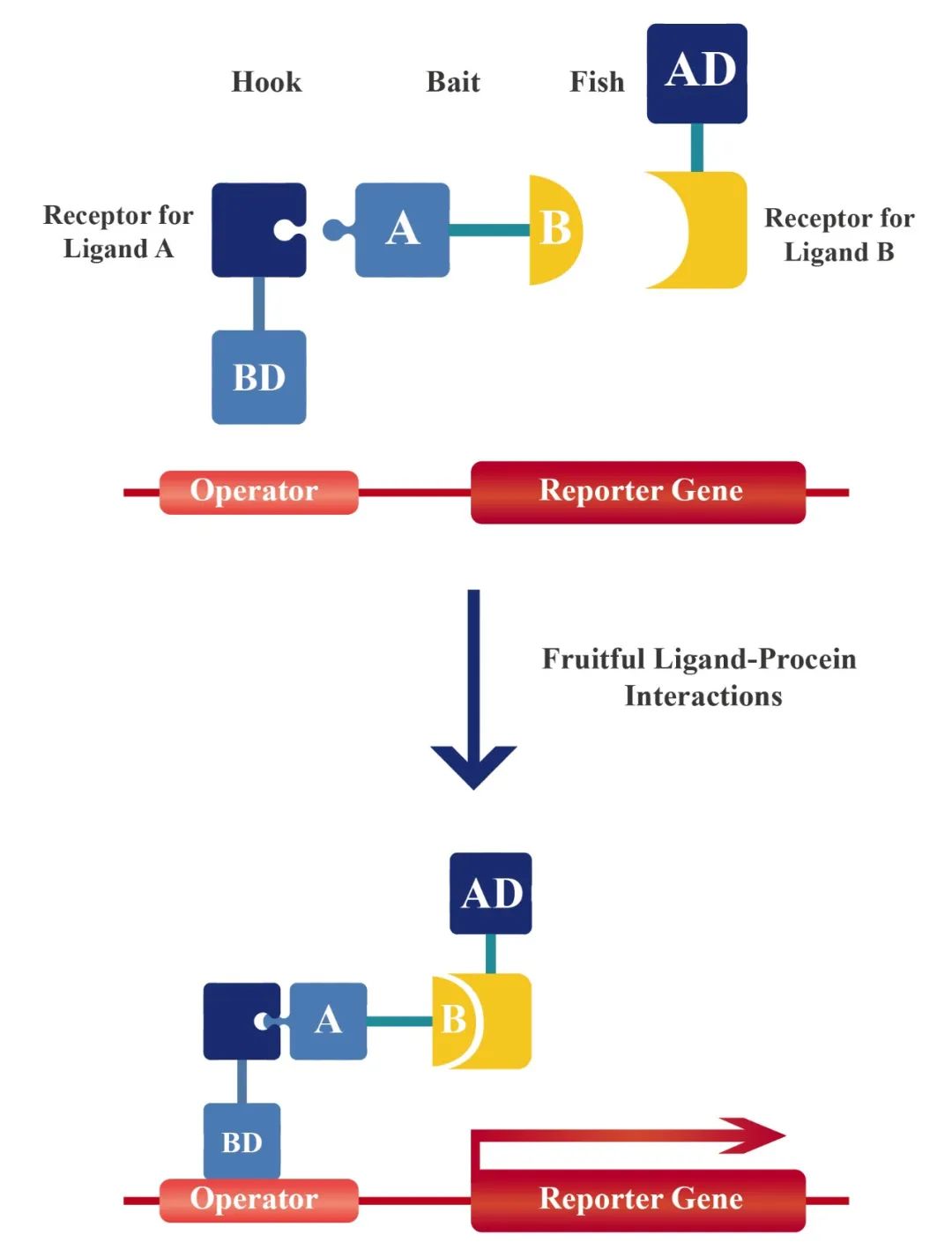
图5 酵母三杂交系统的图解示意图(Licitra Edward J and Jun O Liu., 1996)。合成“诱饵”杂交配体由配体A(正方形)和配体B(半圆形)通过连接剂连接而成。“鱼钩”融合蛋白由配体A受体与转录因子的DNA结合域融合而成。“鱼”融合蛋白由配体B的受体融合到转录因子的反式激活结构域组成。“鱼钩”、“鱼饵”和“鱼”形成一个三聚体复合体,由此使得转录激活域和DNA结合域的重组从而导致报告基因的激活。
应用二
将酵母双杂系统进行简单的改造之后,除了上面讲到的用于研究受体与配体之间的互作之外,还可以用来研究3个蛋白之间的互作,具体信息如下:
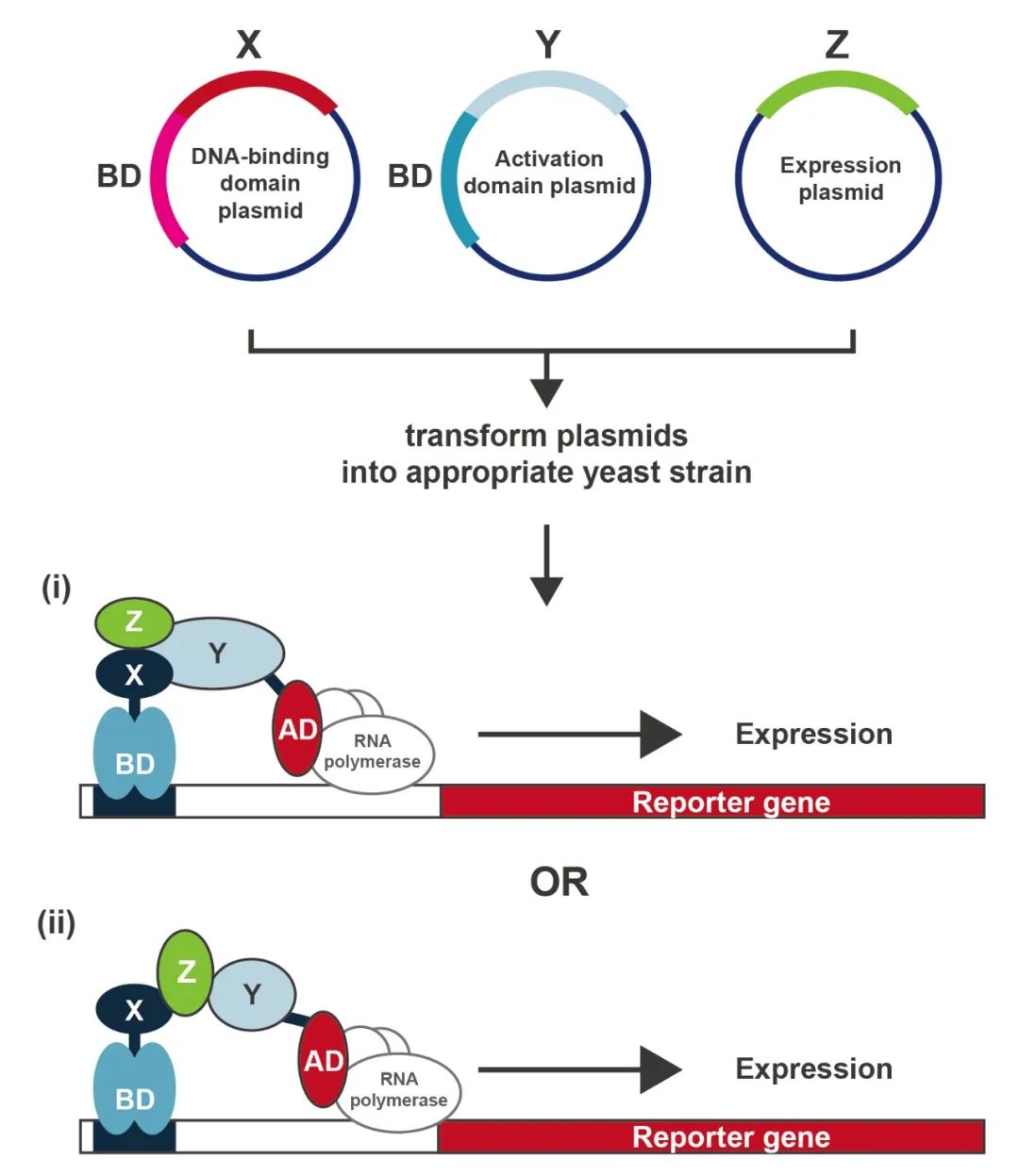
图6 酵母三杂交系统(Causier Barry and Brendan Davies, 2002)。该系统也是基于经典的酵母双杂交系统,蛋白质X和Y分别在具有转录因子DNA结合域和转录激活域的载体内表达。第三种蛋白Z在酵母细胞核中表达,没有添加任何结构域。蛋白质Y可能只在Z存在的情况下与X相互作用。通过X和Z相互作用形成的结构域可能为蛋白质Y(i)提供了一个相互作用界面。或者,蛋白质Z也可以作为蛋白质X和蛋白质Y之间的桥梁(ii)。
应用三
小分子配体在探索蛋白质的细胞功能方面具有不可估量的价值。这些配体的结合导致其目标蛋白的激活或失活,类似于相应基因中功能获得或丧失突变所产生的效应。利用具有细胞渗透性的小分子配体来改变蛋白质的功能是很有吸引力的,因为其固有的条件可以通过添加或移除这种化合物来简单、快速地控制目标蛋白的活性。为了研究目标蛋白对应的小分子配体是什么,在酵母双杂的基础上开发以下方法:
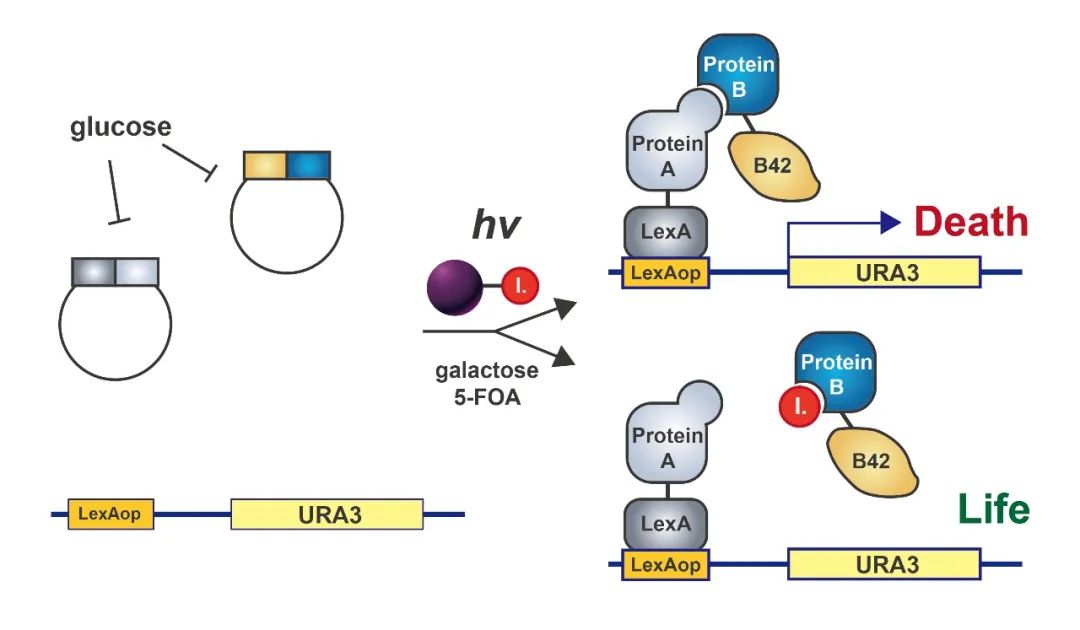
图7 用于检测蛋白质相互作用的小分子抑制剂的反向双杂交系统的示意图(Huang Jing and Stuart L Schreiber, 1997) 。该系统也是基于经典的双杂交系统,但是与经典双杂交系统不一样的是报告基因的表达在一定条件下对酵母细胞是有毒的。相互作用蛋白的表达由GAL1启动子控制,在Glc中被抑制。在Gal诱导后,两个相互作用的蛋白质被合成,它们的结合反过来会诱导合成一种有毒的基因产物,导致细胞死亡,除非在细胞中存在一种蛋白质相互作用的抑制剂。通过这种方式验证抑制蛋白互作的小分子或目标蛋白的小分子配体。
(2)膜体系酵母双杂
传统酵母双杂交系统的各种修饰在确定或检测细胞核和细胞质蛋白之间的蛋白-蛋白相互作用中发挥了重要作用,其原理是基于转录因子(TF)活性的重建,其过程必须发生在细胞核中(Fields Stanley and Ok-kyu Song, 1989)。这种方法可以识别新的互作蛋白,并为它们的功能提供线索。然而,膜蛋白,如受体酪氨酸激酶、G蛋白偶联受体、膜结合磷酸酶和转运蛋白等重要的信号分子,由于其疏水性,很难使用经典的蛋白质相互作用分析方法进行研究。因此,需要寻找另外一种方法对其进行研究,并且该方法允许识别完整的膜相互作用蛋白。对这一块知识比较了解的应该都知道该方法就是膜体系酵母双杂交(MbYTH)。
MbYTH系统是基于观察到的一个现象,即泛素(Ub)可以通过实验被分离成两个基团,当它们靠近彼此时,其功能又可以进行重构 (Johnsson Nils and Alexander Varshavsky, 1994)。因此,分裂泛素的重组就可以用来研究融合到泛素部分的膜蛋白的相互作用(Stagljar et al., 1998)。那么,泛素到底是什么呢?泛素是一种进化保守的,包含76个氨基酸的蛋白,可作为26S蛋白酶体降解目标蛋白的标签。泛素的存在被位于所有真核细胞细胞核和细胞质中的泛素特异性蛋白酶(UBPs)识别。
MbYTH系统中也包括两个载体:(1)将一种感兴趣的膜蛋白,即所谓的诱饵,融合到半个泛素(Cub,即泛素分子被截断后的C端)上,并且Cub上还连有一个人工TF,该TF由细菌LexA-DNA结合域和单纯疱疹病毒VP16激活蛋白组成;(2)将另一种感兴趣的蛋白,即所谓的猎物,它们要么是膜蛋白,要么是细胞质蛋白,融合到半个泛素(Nub,即泛素分子被截断后的N端)上。
Cub与Nub在同一个细胞中存在时能够自发重组形成有活性的泛素,因此将Nub中的第13位亮氨酸突变为甘氨酸或丙氨酸(NubI-NubG或NubA)后,可使它与Cub之间的亲和力降低,从而可以避免这种自发重组。在这些突变体中,只有当分别与NubG/A和Cub融合的两个蛋白间有相互作用时,才能在空间上使NubG/A和Cub足够接近形成可以被UBPs识别的泛素,随后UBPs将融合于Cub的C端末端的TF解离释放。释放的TF随后进入细胞核,激活报告基因如HIS3、LacZ、ADE2的转录与表达,这个可以通过酵母在选择板上的生长或比色法进行监测(图8)。
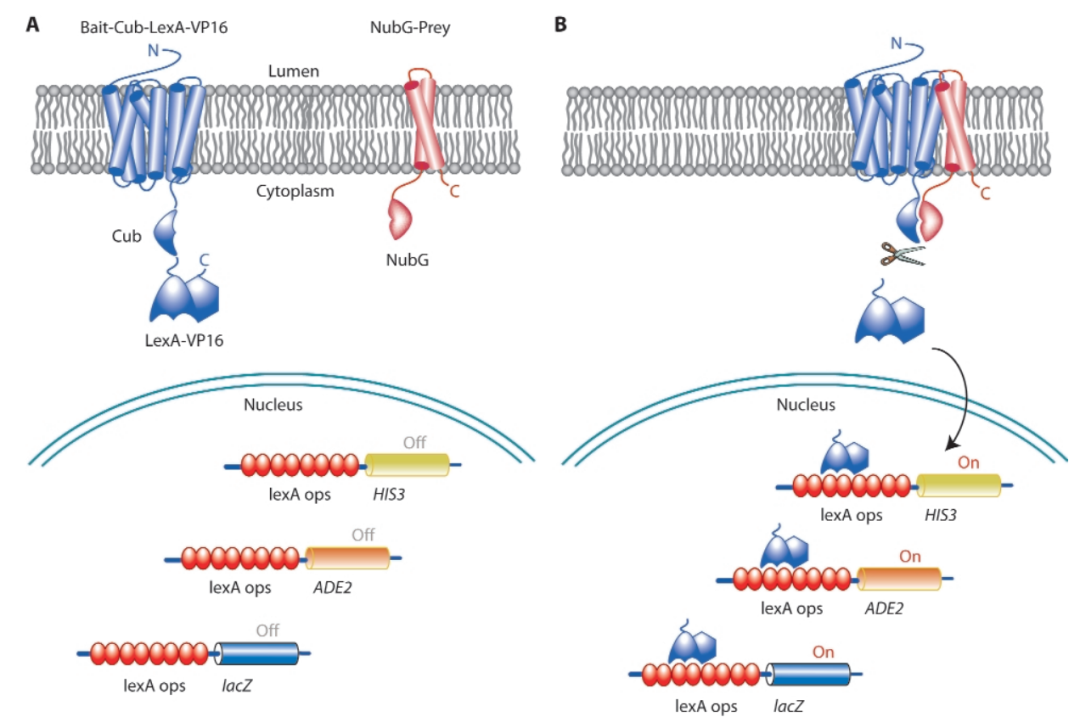
图8 膜体系酵母双杂概述(Iyer et al., 2005)。(A)一个感兴趣的膜诱饵蛋白与Cub融合,然后是人工转录因子LexA-VP16(蓝色),而另一个猎物膜蛋白(或细胞质蛋白)融合到NubG结构域(红色)上。如果诱饵和猎物不存在相互作用,则不会发生泛素重组,也不会发生UBPs介导的转录因子裂解;这导致酵母HIS3-/ADE2-和LacZ-的结果。(B)当诱饵和猎物蛋白相互作用时,泛素发生重组,导致UBPs的蛋白水解裂解和转录因子的释放。该因子进入细胞核,通过与启动子内的Lex A操作位点(lexA ops)结合来激活报告基因。这导致酵母HIS3+/ADE2+和LacZ+的结果。
这种所谓的分裂泛素膜酵母双杂交实验涉及将泛素的一半融合到两个相互作用的蛋白质中,其中至少有一个是膜结合的。在这两种蛋白的相互作用下,切断的泛素被聚集在一起,融合到膜蛋白上的转录因子被裂解并释放。自由转录因子进入细胞核,激活报告基因的转录。
酵母三杂
酵母三杂实验的意义
在细胞中,RNA与蛋白质的相互作用对于包括病毒传播在内的各种生物过程都是至关重要的。例如,在真核生物中,细胞需要大量的RNA结合蛋白来控制其mRNA的含量。mRNA的代谢包括从核内合成到细胞质内降解的一系列过程,每一步都需要大量的RNA结合蛋白和核糖核蛋白(RNPs)。因此,世界各地的分子生物学实验室对RNA-蛋白质相互作用研究的方法提出了越来越多的需求。特别是利用分子遗传学或生物化学方法来识别与之前已确定的RNA目标结合的未知蛋白质,对破译这些基本的生物过程至关重要。在这种情况下诞生了酵母三杂交技术。
酵母三杂原理
1996年,Wickens和Kuhl实验室独立开发了酵母三杂交系统。酵母三杂交系统的总体策略如图9所示,该系统由两个实验室开发,其RNA和蛋白质成分不同(SenGupta et al., 1996;Putz et al., 1996)。该系统的原理是基于酵母细胞报告基因上游RNA-蛋白相互作用介导的多亚基反式激活复合物的组装。这种方法的主要优点之一在于RNA-蛋白质相互作用是在体内进行分析的。反式激活复合物由三个嵌合分子组成。第一个杂交蛋白是与DNA结合域融合的RNA结合蛋白。第二个杂交蛋白分子是一个融合蛋白,其中包含一个融合到Gal4激活域的第二个RNA结合蛋白。第三种杂交分子是一种RNA分子,通过为RNA结合蛋白提供两个特定的RNA靶点来连接两个融合蛋白。
在SenGupta等人1996年开发的方法中,第一个杂交分子是两个蛋白质结构域的融合:大肠杆菌LexA DNA结合结构域(LexA DB),与LexA操作DNA序列和噬菌体MS2涂层蛋白(CP)特异性地结合。为了特异性识别MS2 RNA靶点,MS2 CP需要二聚体化。包含MS2靶标的杂交RNA可以与任何感兴趣的RNA靶标X连接。第二个融合蛋白从一个融合到Gal4激活域的cDNA文库中表达。如果蛋白Y可以与感兴趣的RNA相互作用,这将导致一个活跃的反式激活物复合物的重组,并触发两个报告基因HIS3和LacZ的表达。因此,在活酵母细胞中,通过测量报告基因的表达水平,可以监测RNA X和蛋白Y之间唯一的RNA蛋白相互作用。
在Putz等人开发的方法中,第一个杂交蛋白包含与HIV-1 Rev M10变体融合的Gal4 DNA结合域。杂交RNA分子由Rev RNA靶标组成,即与感兴趣的RNA X相连的Rev反应元件(RRE)。第三个杂交蛋白,Y与Gal4 AD融合,在两个系统中是相同的。
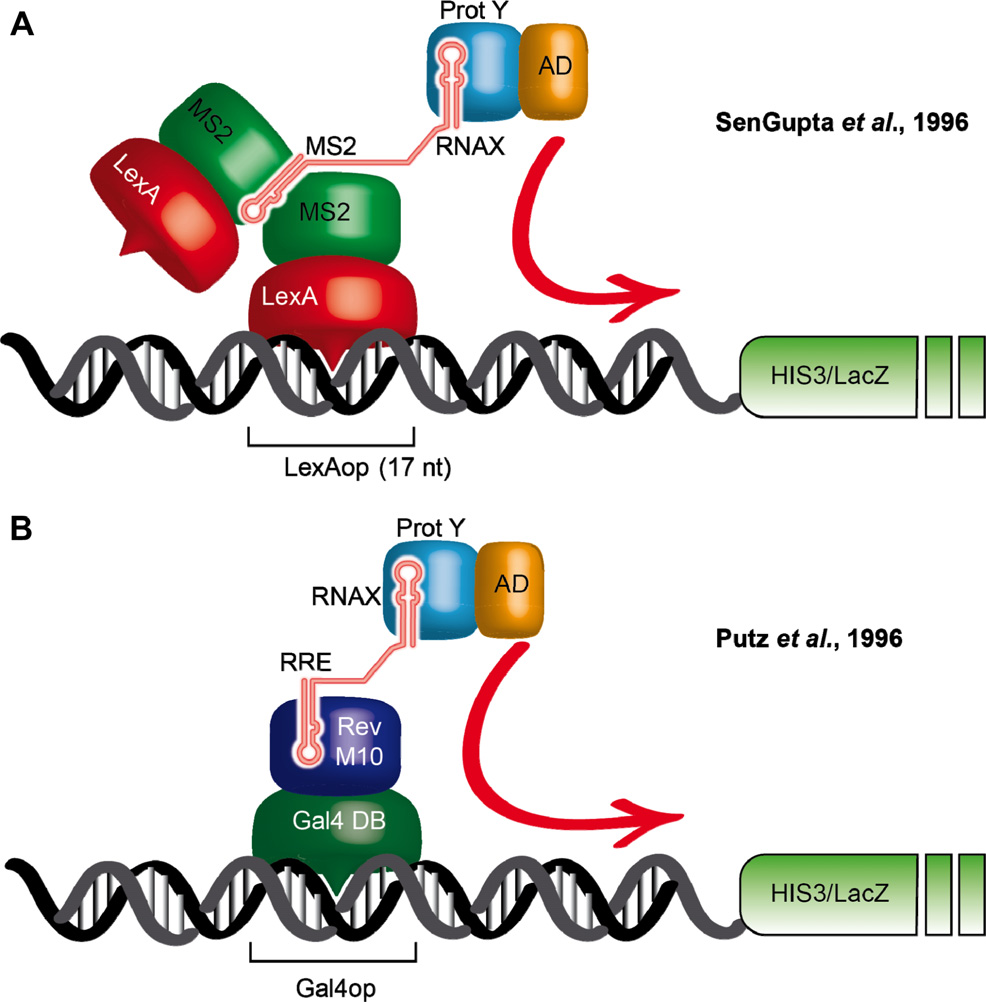
图9 酵母三杂交系统(Brückner et al., 2009)。(A)SenGupta等人设计的酵母三杂交系统(SenGupta et al., 1996)。第一个杂交蛋白包含LexA DNA结合域和噬菌体MS2外壳蛋白(CP)。MS2 CP必须形成二聚体才能与RNA靶点结合。第二个RNA杂交分子包含一个MS2结合位点,该位点可以与任何感兴趣的RNA X相连。第三个杂交蛋白是蛋白质Y与RNA诱饵和Gal4激活域(AD)相互作用的融合蛋白。RNA X与蛋白Y的相互作用导致在17-nt LexA操作子(LexAop)上组装成一个完整的反式激活复合物,该复合物位于两个报告基因HIS3和LacZ的上游。8和4个LexAop分别位于LacZ基因和HIS3基因的上游。(B)在Putz等人1996年的系统中,第一个杂交蛋白由与HIV Rev M10突变蛋白融合的Gal4 DNA结合域组成。该蛋白与其RNA靶标,Rev反应元件(RRE)结合。第三个杂交蛋白和报告基因在两个系统中是相同的。
酵母三杂交筛库体系不仅可以利用RNA作为诱饵去筛选与之互作的蛋白,也可以利用蛋白作为诱饵去筛选与其互作的RNA。另外,由于某些RNA具有激活报告基因转录的能力,这意味着它们不需要任何AD蛋白就能触发报告基因的表达。根据这一特性设计出了一种新型RNA杂交系统用于检测RNA-RNA相互作用,其原理如下:
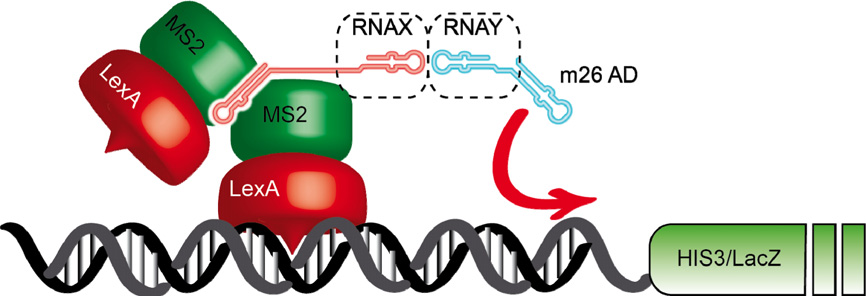
图10 酵母三杂交方法的变体用于检测RNA-RNA相互作用(Brückner et al., 2009)。这个体系包含两个杂交RNA分子。第一种杂交与经典的三杂交方法相同,即MS2与任何感兴趣的RNA X融合。第二种RNA杂交是与自反式激活RNAm26相连的特定RNA相互作用子Y之间的融合。X和Y相互作用导致HIS3和LacZ报告基因的表达。
酵母单杂与酵母双杂是我们经常会用到的实验,想必大家都比较熟悉,但是膜系统酵母双杂与酵母三杂了解的人应该就没那么多了,不管之前如何,今天通过伯远生物(https://plant.biorun.com)细致地讲解,大家对它们的原理应该都比较清楚了!看完文章有不懂的地方可以与伯小远交流哦!另外文章中提到了酵母双杂衍生出来的酵母三杂还有验证RNA与蛋白互作的酵母三杂,很多时候名字都一样,大家一定要区分清楚哦!原理比较复杂,但是理解了就会觉得很简单,希望大家好好吸收,后面将继续为大家讲解酵母杂交相关的知识及具体文献举例,大家记得关注哦!
References:
Bhaumik S R, Raha T, Aiello D P, et al. In vivo target of a transcriptional activator revealed by fluorescence resonance energy transfer[J]. Genes & development, 2004, 18(3): 333-343.
Brückner A, Polge C, Lentze N, et al. Yeast two-hybrid, a powerful tool for systems biology[J]. International journal of molecular sciences, 2009, 10(6): 2763-2788.
Causier B, Davies B. Analysing protein-protein interactions with the yeast two-hybrid system[J]. Plant molecular biology, 2002, 50(6): 855-870.
Fields S, Song O. A novel genetic system to detect protein–protein interactions[J]. Nature, 1989, 340(6230): 245-246.
Huang J, Schreiber S L. A yeast genetic system for selecting small molecule inhibitors of protein–protein interactions in nanodroplets[J]. Proceedings of the National Academy of Sciences, 1997, 94(25): 13396-13401.
Iyer K, Burkle L, Auerbach D, et al. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins[J]. Science's STKE, 2005, 2005(275): pl3-pl3.
Johnsson N, Varshavsky A. Split ubiquitin as a sensor of protein interactions in vivo[J]. Proceedings of the National Academy of Sciences, 1994, 91(22): 10340-10344.
Keegan L, Gill G, Ptashne M. Separation of DNA binding from the transcription-activating function of a eukaryotic regulatory protein[J]. Science, 1986, 231(4739): 699-704.
Licitra E J, Liu J O. A three-hybrid system for detecting small ligand–protein receptor interactions[J]. Proceedings of the National Academy of Sciences, 1996, 93(23): 12817-12821.
Melcher K, Johnston S A. GAL4 interacts with TATA-binding protein and coactivators[J]. Molecular and Cellular Biology, 1995, 15(5): 2839-2848.
Ouwerkerk P B F, Meijer A H. Yeast one‐hybrid screening for DNA‐protein interactions[J]. Current protocols in molecular biology, 2001, 55(1): 12.12. 1-12.12. 12.
Putz U, Skehel P, Kuhl D. A tri-hybrid system for the analysis and detection of RNA-protein interactions[J]. Nucleic acids research, 1996, 24(23): 4838-4840.
SenGupta D J, Zhang B, Kraemer B, et al. A three-hybrid system to detect RNA-protein interactions in vivo[J]. Proceedings of the national Academy of Sciences, 1996, 93(16): 8496-8501.
Sewell J A, Fuxman Bass J I. Options and considerations when using a yeast one-hybrid system[M]//Two-Hybrid Systems. Humana Press, New York, NY, 2018: 119-130.
Stagljar I, Korostensky C, Johnsson N, et al. A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo[J]. Proceedings of the National Academy of Sciences, 1998, 95(9): 5187-5192.
Stynen B, Tournu H, Tavernier J, et al. Diversity in genetic in vivo methods for protein-protein interaction studies: from the yeast two-hybrid system to the mammalian split-luciferase system[J]. Microbiology and molecular biology reviews, 2012, 76(2): 331-382.
Wu Y, Reece R J, Ptashne M. Quantitation of putative activator‐target affinities predicts transcriptional activating potentials[J]. The EMBO journal, 1996, 15(15): 3951-3963.